

Занимаясь интродукцией растений, невозможно остаться в стороне от селекции. Процесс выращивания тех или иных растений в новой для них среде сам по себе является непроизвольной селекцией, происходит осознанный либо неосознанный отбор наиболее приспособленных экземпляров, и локальное "направление эволюции" вида сменяется вместе со "сменой места жительства". И хотя растения Сада леса ещё маленькие, пора разработать базовые схемы и подходы к дальнейшей работе с ними, так как от этих схем зависит так же и определение мест для высадки растений на территории, и некоторые агротехнические приемы. И даже в самом питомнике может и должен начинаться первоначальный отбор.
Подходы к селекции сильно отличаются в зависимости от ставящихся целей, от физиологических особенностей видов, от места интродукции. Я решила сделать небольшой конспект на память о селекции древесных, а затем разработать несколько базовых схем, разделив растения на группы. Базовые схемы можно будет в дальнейшем корректировать для каждого вида, но они значительно упростят работу на первых этапах. Итак, конспект сделан на основе учебника А. П. Царев, С. П. Погиба, В. В. Тренин "Селекция и репродукция лесных древесных пород".
При селекционном процессе используют методы и приемы разных смежных наук, но основные методы следующие: отбор, гибридизация с использованием гетерозиса и цитоплазматической мужской стерильности, полиплоидия, мутагенез и др. При выборе подходящих методов и схем важно сразу определиться с целями: проводится ли селекция на устойчивость к климатическим условиям или болезням и вредителям, на скорость роста, на качество древесины или семян, на урожайность и качества плодов, на содержание в тканях растений тех или иных хозяйственно-ценных веществ.
Программа селекции для каждого отдельного вида должна включать в себя:
- разработку модели "идеального сорта" (цель программы);
- разработку метода селекции, его реализацию, получение новых форм;
- изучение и первичное испытание полученного потомства и отбор представителей, наиболее полно отвечающих поставленным целям, включая испытание в разных экологических условиях.
Эти пункты могут и должны быть пройдены по кругу многократно, прежде чем работу можно будет считать условно завершенной, впрочем, разве может селекция (как и эволюция) считаться окончательно завершенной, особенно в меняющемся мире Земли?
Методология составления программы требует прохождения нескольких шагов.
Шаг первый. Модель сорта
При всем различии целей, все сорта лесных древесных должны иметь следующие признаки:
- высокую продуктивность по селектируемому признаку в условиях данного региона;
- высокую экологическую стабильность для возможно более широкой амплитуды экологических условий;
- высокую зимостойкость, а также устойчивость к почвенной и атмосферной засухе;
- повышенную устойчивость к ветру, снеговалу и снеголому;
- устойчивость к болезням (корневой и стволовой гнили, ржавчине, фузариозу, раковым заболеваниям и т. д.), а также к энтомовредителям.
Для конкретных целей список должен быть дополнен специфическими требованиями. Все эти требования и являются каркасом для модели "идеального сорта".
Шаг второй. Изучение исходного материала
Следующим важным шагом будет изучение исходного материала: внутривидовой и видовой потенциал, как искусственный (сорта, гибриды, мутанты и т. д.), так и естественный (разновидности, расы, формы).
Изменчивость живых существ является основой органической эволюции. Изменчивость делится на качественную и количественную. Качественные признаки обычно имеют дискретное значение (опушенный/голый, гладкий/морщинистый), чаще всего контролируются одним или немногими генами и наследуются в соответствии с законами Г. Менделя. Количественные признаки являются суммарным результатом действия многих генов, поддаются подсчетам и измерениям и обладают непрерывной изменчивостью. Проявление количественных признаков сильно зависит от условий среды.
Внутривидовая изменчивость подразумевает, что растения, принадлежащие к одному виду, можно делить на несколько типов категорий. По В. Н. Сукачеву это:
- экотипы (климатипы, эдафотипы, ценотипы) - результат групповой изменчивости;
- лузусы - формы, отличающиеся каким-то признаком, но не обладающие особым ареалом и не принадлежащие к определенным экотипам, а могущие встречаться в разных ареалах и условиях среды;
- аберрации - редкие резко уклоняющиеся формы;
- экады, модификации - ненаследственные формы.
Большое значение имеет закон гомологических рядов, сформулированный Н. И. Вавиловым. Разнообразие форм одного вида или рода можно прогнозировать и у родственных ему видов и родов.
Изучение изменчивости исходных популяций позволяет выбрать наиболее эффективный метод селекции.
При наблюдении за изменчивостью растений важно разграничить влияние среды и наследственности на формирование конкретного фенотипа. Гетерогенность популяций наиболее значительна в области оптимума вида и ниже в экстремальных условиях, однако в экстремальных условиях бывает повышена частота отдельных форм. В изменении отдельных групп признаков могут наблюдаться корреляции.
Чем лучше будет изучена внутривидовая изменчивость, её механизмы и особенности в каждом конкретном случае, тем более удачная программа селекции может быть составлена.
Шаг третий. Выбор метода селекции
1) Отбор
Основным приемом в селекции является отбор. Отбор делится на естественный (стабилизирующий, дизруптивный, направленный) и искусственный (бессознательный, либо же методический).
Методический отбор может быть прямым (по селективному признаку) и непрямым (по косвенному признаку, связанному с селективным). Методический отбор делится на массовый и индивидуальный.
Массовый отбор
В основе массового отбора лежит оценка фенотипа, по которому выделяют множество индивидов и размножают их совместно. Это самый простой вид отбора, однако его недостатком является трудность учета влияния среды на фенотип, а также невозможность добиться результата, если популяция, в которой ведется отбор, гомозиготна по генам, определяющим селектируемый признак. При проведении массового отбора по фенотипу в гетерозиготной популяции перекрестников изменение признака в желаемую сторону идет быстро в первых поколениях, а затем замедляется по мере того, как отбираемые особи становятся более гомозиготными по желательным генам. Чем большее число особей участвует в отборе, тем больше длится эффективный массовый отбор.
Выбор желательных типов в популяции называется позитивным отбором, а выбраковка нежелательных - негативным. При позитивном массовом отборе в лесном хозяйстве применяются разные методы:
- отбор лучших климатипов (семена для выращивания должны быть взяты либо местные, либо из схожего климата, из более подходящих частей природного ареала вида, для основных пород деревьев были проведены множественные масштабные исследования, которые можно найти, например, в специальном издании "Лесосеменное районирование основных лесообразующих пород в СССР");
- отбор лучших эдафотипов в пределах какого-то географического района (при подборе семян для посева надо учитывать не только климат места произрастания родительских деревьев, но и почвенные условия, тип леса);
- отбор лучших (плюсовых) насаждений ("групповой отбор" - для заготовки семян используется лучшее насаждение, выбранное в данном географическом или лесосеменном районе, эффективен для признаков, передающихся наследственно и явно отражающихся в фенотипе, таких как устойчивость к снеголому);
- отбор лучших (плюсовых) деревьев (отбираются лучшие деревья, из черенков которых создают прививочные семенные плантации первого порядка);
- отбор лучших сеянцев в питомнике и семян по крупности и качеству (в основном, проводится ради получения растений с более энергичным ростом в первые годы жизни).
Итого, массовый отбор может давать неплохие результаты при отборе на улучшение формы, на устойчивость к болезням. Эффективность массового отбора зависит от наследуемости признака, размера популяции, её гетерогенности. Недостатками массового отбора считается:
-невозможность проверки отбираемых растений по потомству (в общем сборе семян, наряду с семенами наследственно ценных деревьев, могут попасться семена с деревьев, которые кажутся ценными в результате положительных модификаций под действием благоприятных условий);
-невозможность выделить наиболее ценные формы, их особые качества не скажутся на среднем значении, по которому подбирались источники;
-отбор генотипов только по материнской линии;
-при малом числе отбираемых растений в результате генетического дрейфа или инбридинга может привести к ухудшению сорта.
Индивидуальный отбор
Отбирают отдельные растения, потомство каждого из которых размножают в дальнейшем отдельно. Отобранные растения многократно проверяются по потомству, неудачно отобранные растения, что проявится в худшем потомстве, выбраковываются вместе с потомством. Основные методы индивидуального отбора: метод педигри (для самоопыляющихся растений), клоновый отбор (для вегетативно размножаемых), отбор у перекрестноопыляющихся растений.
Метод педигри
Используется при селекции самоопыляющихся растений. Отобранные растения высеваются индивидуально (линиями), в каждом следующем потомстве выбраковывают худшие линии, а в оставшихся линиях проводят индивидуальный отбор лучших растений и от них высевают следующее поколение, после чего цикл повторяется с новым поколением. После того как линии достигнут высокой константности (в F5-F6), растения внутри них объединяют и лучшие линии высевают для сравнительного испытания на соответствующих площадях.
Клоновый отбор
Индивидуальный отбор в селекции растений, размножаемых вегетативно. Отбирают лучшие растения, размножают вегетативно, и полученное потомство от нескольких выбранных растений сравнивают в разных условиях друг с другом (позволяет корректно тестировать одни и те же генотипы в разных условиях).
Индивидуальный отбор у перекрестноопыляющихся растений
Для перекрестноопыляющихся растений вследствие постоянного переопыления и расщепления признаков сорта нельзя вывести путем однократного отбора, а применяют непрерывный индивидуальный отбор, при этом имеют дело не с отдельными растениями, а с семьями, то есть с потомством одного растения. Из семян отдельных растений, принадлежащих семье, высевают, образуя новую дочернюю семью. Семьи сравнивают между собой, худшие бракуют. Смысл метода в повторении отбора лучших растений из лучший семей. Так как у перекрестноопыляемых растений требуется следить за наследственностью отцовских растений, метод делится на два основных варианта: индивидуально-семейный и семейно-групповой отбор.
При индивидуально-семейном отборе каждая семья размещается изолированно от остальных потомств. По сравнению с первым отбором в исходном материале, второй отбор имеет то преимущество, что отбранные лучшие растения, высаженные изолированной семьей, опыляются пыльцой только в пределах данной семьи показавшей лучшие качества (отрицательной стороной может быть некоторая инцухт-депрессия). Потомства лучших растений второго отбора снова оценивают при изолированном их размещении по семьям, а отбор ведут по потомствам. В конечном итоге лучшие растения последних потомств из разных семей высаживают вместе и размножают совместно, чтобы преодолеть негативные последствия близкородственного скрещивания.
При семейно-групповом отборе из отобранных в исходной популяции растений формируют несколько групп по физиологическим или морфологическим признакам, а потомство лучших растений из каждой группы высаживают изолированно. Внутри группы отбирают лучшие потомства, а остальные выбраковывают. Эти потомства переопыляются между собой в пределах данной группы. Таким образом, вероятность близкородственного размножения уменьшается, а переопыление происходит в рамках сходных семей. Лучшие потомства каждой группы объединяют по принципу массового отбора, смеси из лучших потомств каждой группы испытывают и размножают, в ряде случаев в пределах группы отбор проводят дважды. При таком варианте отбора сохраняются достоинства направленного переопыления, но снижается инцухт-депрессия, хотя выровненность материала достигается медленнее.
Оба варианта индивидуального отбора у перекрестно-опыляемых растений для лесных пород требует 70-100 лет и более.

2) Гибридизация
Гибридизация - получение новых комбинаций генов (но не новых генов!). В современном понимании, гибрид - это гетерозиготная особь, возникающая в результате скрещивания генетически различных родительских генотипов, т. е. любой гетерозиготный организм, независимо от его происхождения. Почти любое скрещивание между лесными деревьями даже одного вида будет производить гибриды. Половая гибридизация является распространенным методом синтетической селекции, она может быть спонтанной или контролируемой.
Цели гибридизации:
- повышение устойчивости растений к негативным факторам среды, увеличение их жизнестойкости и долговечности;
- увеличение быстроты роста и мощности;
- повышение декоративных качеств;
- повышение урожайности и качества плодов, содержания смол и ценных веществ.
- оценка наследуемости;
- оценка общей комбинационной способности (ОКС);
- оценка специфической комбинационной способности (СКС);
- получение исходного материала для селекции;
- оценка генетического выигрыша.
Внутривидовые и скрещивания между близкими видами называют конгруэнтными, или совместимыми, между более дальними видами или близкими родами - инконгруэнтными, или несовместимыми.
Также скрещивания делят на три вида: комбинационные, трансгрессивные и гетерозисные.
Комбинационные скрещивания
Используются для объединения двух или нескольких полезных признаков в одном растении. Установлены некоторые закономерности формообразовательных процессов в гибридных популяциях и разработаны принципы подбора родительских пар (эколого-географический, по элементам структуры урожая, по продолжительности отдельных фаз вегетации, по устойчивости к заболеваниям)
При любых подходах при скрещиваниях получается широкий спектр изменчивости в гибридном потомстве.
Трансгрессивные скрещивания
Цель - получить трасгрессию, эффект суммирующего действия полимерных генов, в результате которого устойчиво увеличивается или уменьшается значение количественного признака у отдельных гибридных особей потомства F2 по сравнению с крайними значениями этого признака у родительских форм. Семенное потомство полученных особей в результате самоопыления или опыления между собой будет так же трансгрессивно.
Гетерозисные скрещивания
Гетерозис истинный, если гибрид превосходит по признаку лучшего родителя и гипотетический, если превосходит по среднему значению признака у родителя. Также гетерозис может быть соматическим (мощное развитие вегетативных органов), репродуктивным (высокий урожай и фертильность), адаптивным (повышенная приспособленность).
Принципиальное практическое отличие гетерозиса от трансгрессии в том, что он проявляется только у F1.
Категории гетерозиса: популяционный (в результате длительной адаптивной эволюции в панмиктических естественных популяциях, в результате естественного отбора, поддерживающего гетерозиготы), групповой (при гибридизации форм с высокой СКС, когда семья в целом по средним и максимальным показателем превосходит обе родительские формы) и индивидуальный (только отдельные особи гибридной семьи превосходят родительские формы).
Наиболее подходящие методы гибридизации зависят от физиологических особенностей вида, требований к будущему сорту и т. д.
Простые скрещивания
Однократное скрещивание между двумя родителями. Разновидность простых скрещиваний - реципрокные скрещивания, когда параллельно скрещиваются две родительских формы, но в одном случае первый родитель выбран как материнское растение, а второй как отцовское, а во втором случае наоборот.
Сложные скрещивания
Делятся на ступенчатые и возвратные.
Ступенчатые используются для соединения более чем двух разных родительских форм, производятся последовательно.
- [(A*B)*C]*D;
- [(A*B)*(C*D)]*E.
Возвратные - при которых гибрид скрещивается повторно с одним из родителей, используются:
- для преодоления бесплодия гибридов первого поколения при отдаленной гибридизации (A*B)*B, для усиления в гибридном потомстве;
- для усиления в гибридном потомстве желаемых свойств одного из родителей - насыщающие скрещивания, (А*В)*В*В*В*В*В, каждое следующее скрещивание с одним из родителей называют беккроссом (требуется для выведения ценных сортов с устойчивостью к болезням, тогда А - малоценный устойчивый родитель, а В - неустойчивый, но высокоценный родитель).
Инцухт-гетерозисные скрещивания
При этом методе вначале получают самоопыленные инцухт-линии (инбред-линии), представляющие собой потомство неоднократно принудительно самоопыленного перекрестноопыляющегося растения. Затем отбирают из них инбред-линии с высокой комбинационной способностью и скрещивают между собой. В результате получаются гибриды с повышенным гетерозисом, инцухт-гетерозисные.
Проведение контролируемых скрещиваний лесных деревьев - дорогое и долговременное мероприятие, поэтому важно заранее продумать оптимальный план скрещиваний. Есть несколько принятых схем контролируемых скрещиваний: полную схему диаллельных скрещиваний, модифицированную схему диаллельных скрещиваний, частичные схемы диаллельных скрещиваний, факториальную схему скрещиваний, простые скрещивания.
Полная схема диаллельных скрещиваний - наиболее полная схема, дающая почти всю информацию о наследственных свойствах каждого проверяемого растения:
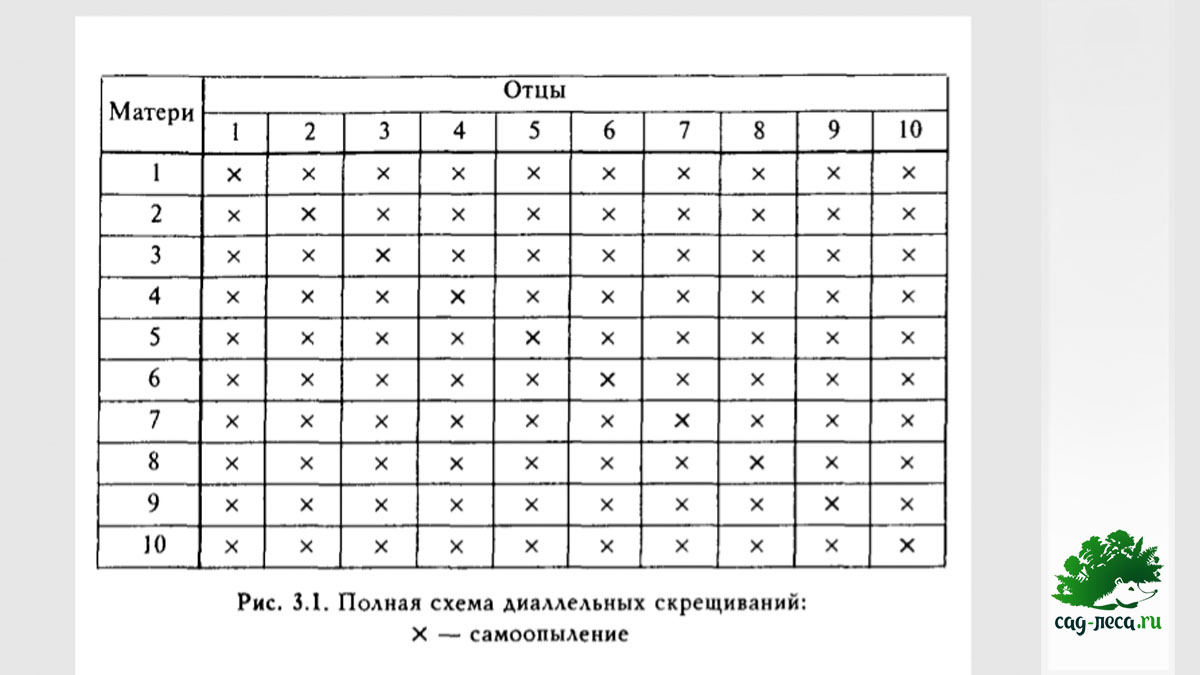
Но она является очень дорогой и трудоемкой, ведь даже небольшое число скрещиваемых экземпляров, возведенное в квадрат, даёт огромное число необходимых контролируемых скрещиваний. Немногим меньше данных, зато намного менее трудоемка модифицированная схема диаллельных скрещиваний, из которой исключены самоопыления и реципрокные скрещивания:
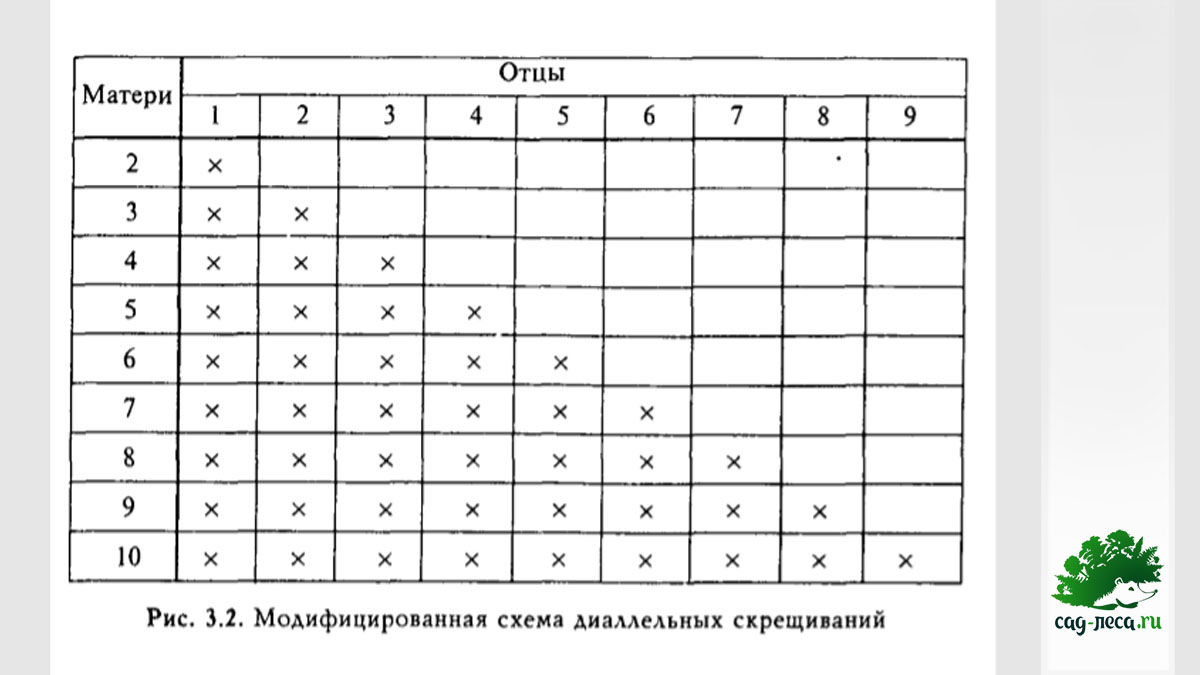
Еще меньше информации дает частичная схема, но она позволяет опробовать большое количество экземпляров относительно недорого:
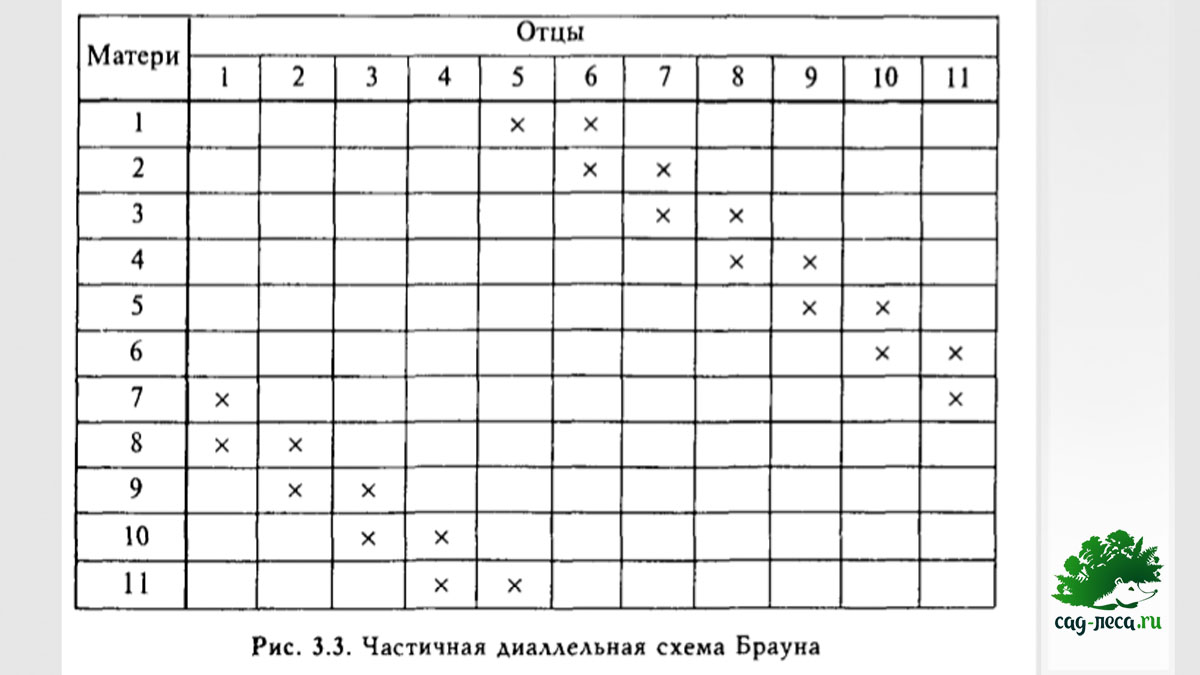
К сожалению, нехватка данных, которые дает частичная схема, значительно усложняет расчеты.
Факториальная схема - это схема, при которой разные материнские клоны скрещиваются с небольшим количеством одних и тех же отцов, которые называются общими тестерами. Так как одни и те же клоны не используются и как матери, и как отцы, трудно сравнить ОКС каждого родителя.
При схеме простого скрещивания пар каждый экземпляр включен только один раз - как мать или как отец.
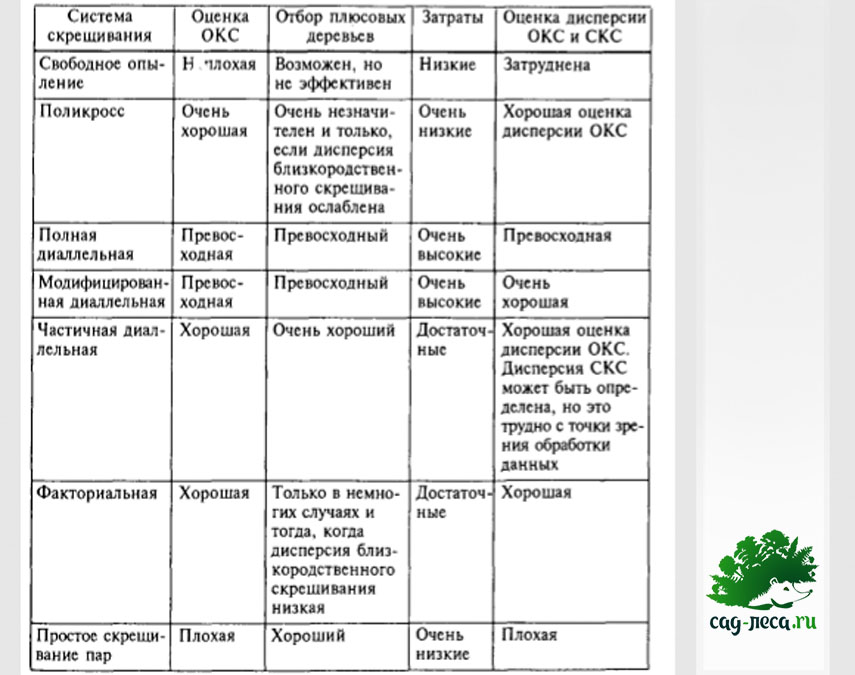
У каждой схемы есть достоинства и недостатки, а также ситуации, в которых они наиболее применимы. Краткое сравнение дано в таблице:
Цветки на отобранных материнских растениях за 1-2 недели до скрещивания изолируются и кастрируются (кроме двудомных видов и тех однодомных видов, у которых разнополые цветки пространственно разобщены). Пыльцу, необходимую для гибридизации, заготавливают заблаговременно за несколько дней до цветения (если используется свежесобранная пыльца). Крупные мужские соцветия в почти зрелом возрасте обрывают и помещают в помещении в бумажные пакеты. Когда пыльца высыпется, её подсушивают в течение часа и собирают в специальные емкости. Хранят пыльцу в недозаполненных пробирках, верх которых закрыт ватой, в холодильнике, в эксикаторе с хлористым кальцием.
Если сроки цветения мужских особей позднее, ветви заранее устанавливают в теплое помещение в воду с питательным раствором, который регулярно меняют, а срезы обновляют.
Опыление производится либо мягкой кисточкой, либо запуская пыльцу в изолятор к цветкам через дырочку, которую потом заклеивают. Опыление производят дважды с интервалом 1-2 дня. Изоляционные пакеты снимаются после появления завязей.
Гибридизация в селекции лесных древесных культур имеет наибольший смысл, прежде всего, для получения ценных генотипов вегетативно размножаемых растений.